|

La simbiogenesi
Un breve profilo storico
Bisogna dire che usare il termine
endosimbiosi (dal greco: ἔνδον: dentro; συν: insieme; βιος: vita), come equivalente di
simbiogenesi, è errato. Endosimbiosi è termine solo adeguato a ciò che in natura si pone nel campo ricco della simbiosi, che produce vantaggi per entrambi i soggetti in causa o solo per uno (mutualismo, commensalismo, parassitismo), nelle localizzazioni extracellulari o intracellulari degli endosimbionti. La simbiogenesi, invece, si vuole porre come fondamento dell’evoluzionismo.
Si è convenuto, nel
vulgato, di attribuire l’ipotesi della simbiogenesi a Lynn Margulis, americana di famiglia ebraica (1938 - 2011), ma la biologa andrebbe, con più ragione, ricordata per la sua opera divulgatrice della simbiogenesi. Infatti, alle spalle dell’opera di Lynn Margulis c’è una storia di idee, alle quali è molto debitrice.
La sua opera di diffusione si è svolta dal 1966 con un primo articolo: “The origin of mitosing Eukaryotic Cell”, pubblicato dal “Juornal of Theoretical Biology”. A questa pubblicazione seguì nel 1970 il libro. “Origin of Eukaryotic Cell”, edito da Yaale Universiy Press. Poi, nel 1981 uscì il libro: “Symbiosis in Cell Evolution”, W, H. Freeman and Company, New York. La produzione letteraria di Lynn Margulis assomma a una dozzina di libri, alcuni scritti con la collaborazione del figlio Dorian Sagan; inoltre si hanno circa 130 articoli. Nel 1983 venne nominata membro della
National Academy of Science.
L’idea e la parola “Simbiogenesi” sono state introdotte dal botanico russo Konstantin Sergeevic Merezkovski (1905), la cui idea ebbe uno sviluppo in Russia e nell’Unione Sovietica, che solo di recente è venuta a conoscenza degli scienziati occidentali con una pubblicazione di Liya Nikolaevna Khakhina (1979): “Historical and Critical Study of the Research of Russian Botanists”, con prefazione di Lynn Margulis e Mark McMenamin. Il testo venne pubblicato in Russia nel 1979.
Merezkovski era al corrente delle acquisizioni del botanico tedesco Andreas Schimper, che, nel 1883, con un microscopio aveva visto come la divisione dei
cloroplasti, propri delle cellule vegetali, ricordava la divisione dei Cianobatteri, da ciò aveva congetturato che ci fosse stata un tempo un’unione simbiotica, poi fissatasi, tra due organismi.
Nel 1909 lo zoologo Umberto Pierantoni avanzò il pensiero di una simbiosi fisiologica ereditaria.
Nel 1918 il francese Paul Jules Portier avanzò l’idea che i mitocondri provenivano da un ancestrale processo di simbiosi.
Nel 1920 il biologo americano Ivan Emanuel Wallin avanzò la medesima ipotesi circa i mitocondri, fondandosi su culture di mitocondri isolati dalle cellule di fegato fetale di coniglio, in apposite soluzioni. La moltiplicazione dei mitocondri era però solo una sua idea poichè la grande maggioranza delle loro proteine (circa il 95%) sono prodotte sulla base del DNA nucleare della cellula, solo il restante 5% delle proteine proviene dal DNA mitocondriale. I contemporanei rifiutarono gli esperimenti del Wallin dicendo della presenza di contaminanti batterici, e tutto finì.
Grande pioniere della microscopia elettronica fu Hans Ris (1914 - 2004), di origini svizzere. Lynn Margulis studiò presso l’Università del Wisconsis (1960), dove ebbe l’opportunità di conoscere Hans Ris. Hans Ris vide col microscopio che all’interno dei cloroplasti delle cellule vegetali, dove si compie la sintesi clorofilliana, c’era del DNA a cerchio come nei batteri, e i cloroplasti si moltiplicavano all’interrno della cellula in modo autonomo dalla stessa e per scissione binaria, come per i batteri. L’autonomia dei cloroplasti ha dei limiti poiché la grande maggioranza delle sue proteine - come per il mitocondrio - non sono codificate dal suo DNA, ma da quello del nucleo della cellula: prodotte nel citosol (citoplasma) e poi immesse nel cloroplasto.
Una simbiogenesi dai batteri è solo e unicamente affidata alla fantasia.
La microscopia elettronica permise anche di vedere che i mitocondri avevano due membrane, similmente ai batteri.
L’analisi della membrana interna dei mitocondri ha portato a rilevare che un quinto della loro composizione lipidica è costituito dalla
cardiolipina (scoperta negli anni 40), che si ritrova nelle membrane, con proprie percentuali, della maggior parte dei batteri. La
cardiolipina è stata assunta come una prova ipotetica della simbiogenesi. La
cardiopilina (difosfatidil-glicerolo), tuttavia, ha un potenziale di complessità enorme, per il quale varia le sue configurazioni strutturali, e il suo apporto, nel contesto delle varie realtà biologiche.
Hans Ris considerò l’ipotesi della simbiogenesi.
Lo specialista in microscopia elettronica è stato, insieme a Lynn Margulis, uno dei primi ad affermare che le alghe blu-verdi erano un tipo speciale di batteri, i
Cyanobacteria, un philum di batteri fotosintetici.
La posizione metafisica e religiosa di Lynn Margulis
Lynn Margulis ha professato un vago deismo estremista. “Penso che ciascuno possa credere quello che vuole”: (Intervista di Paolo Musso: marzo 2009). “Ammetto le mie convinzioni: tutte le religioni organizzate sono inganni istituzionalizzati, imbrogli condivisi e selvaggio e derisivo tribalismo”: Lynn Margulis: (“Una revolucion en la evolution: escritos seleccionados”; Universitat de Valencia, 2002, pag. 45-48).
Lynn Margulis ebbe certo una complicità di pensiero religioso con il primo marito Carl Edward Sagan, che dopo alcuni mesi di fidanzamento, sposò (1957). Sagan era un astronomo di valore, ma con indulgenze fantascientiche. Deista e spregiatore delle religioni.
La Margilus anche se rigettava, giustamente, il neodarwinismo, che voleva che l’evoluzione fosse avvenuta per accumulo di microevoluzioni, non aderì all’Intelligent design,
perché dichiarava un’Intelligenza creatrice.
Va notato che l’Intelligent design non parte da un’idea a tavolino, ma dall’evidenza della
complessità della cellula, della sua unità non meccanicistica, ma vitalista in se stessa.
L’organellogenesi
Essa postula per la formazione dei cloroplasti che (circa 1,2 miliardi di anni fa), un batterio pre-eucariota, ameboide, (ipotetico prodotto di endosimbiosi parassitarie tra procarioti) abbia fagocitato un Cianobatterio, e non attuando il processo enzimatico digestivo, causalmente sospeso, sia entrato in endosimbiosi con il
Cianobatterio (alga blu-verde: un procariote), privandolo della parete cellulare, mentre conservava la sottostante membrana. Con il tempo si configurò un cloroplasto, elemento fotosintetico delle alghe verdi, ecc. Da tale processo il cloroplasto avrebbe acquisito una nuova membrana dalla cellula ospite, il batterio pre-eucariota ameboide, il quale avrebbe ricevuto nel suo DNA dei geni del Cianobatterio, mutandosi. Lo stesso è postulato per le alghe rosse, che sono tali perché contengono la ficoeritrina (pigmento rosso) che maschera il cloroplasto verde. Il
vuoto scientifico, che immediatamente si rileva, è l’assenza di spiegazione di come sia avvenuta l’estinzione dello stato di
endosimbiosi per acquisire lo stato di formazione di una nuova entità, cioè la
simbiogenesi. Qui la spiegazione è glissata da un tocco della fantasia.
La simbiogenesi sarebbe passata attraverso la endosimbiosi primaria, segnalata dalle due membrane del cloroplasto. La
endosimbiosi secondaria sarebbe segnalata da tre membrane del cloroplasto, frutto di una fagocitosi, a enzimi digestivi casualmente sospesi, da parte di una supposta
ameba ancestale di un’alga verde o rossa. Le alghe brune, le
alghe dorate, le alghe euglenofite, l’acetabularia mediterranea, hanno cloroblasti a tre o quattro membrane.
Il numero delle membrane dei cloroplasti, per l’ipotesi simbiogenetica è l’indice delle operazioni di fagocitosi.
Ma seguendo la capacità di fotosintesi delle varie alghe alle varie profondità, con intensità luminose diverse e lunghezze d’onda più o meno penetranti il mezzo liquido, il numero delle membrane è funzionale alle loro specifica realtà.
Per la formazione dei mitocondri si ipotizza un batterio pre-eucariota ameboide, fagocitante un piccolo batterio, proposto come
anaerobico, il quale, non venne digerito per errore, e a contatto con le accumulate risorse di ossigeno dell’atmosfera si evolse in mitocondrio.
Si ipotizza che il piccolo batterio appartenesse agli Alphaproteobacteria, in particolare il
SAR11clade, o ai Rickettsiales o, recentemente (2015) alla famiglia degli
Archea: il Lokiarchaeum. Il batterio Lokiarchaeum è stato scoperto presso un’emissione termale nell’Oceano Artico a 5000 m. di profondità. Sarebbero i procarioti più simili agli eucarioti, fino ad ora trovati. Si sottolinea, tuttavia, che gli organismi intermedi alle cellule eucariote non sono ritrovabili, né nel passato né nel presente. E’ poi improbabile che delle proteine di
Lokiarchaeum, “simili” a quelle eucariote, svolgano nel batterio la medesima funzione, non avendo rilevato alcuni enzimi necessari al funzionamento della cellula eucariota (Gautam Dey e Buzz Baum, dell’University College di Londra, e Mukund Thattai del National Centre for Biological a Bangalore, in India).
Si pone l’osservazione che, posto un errore digestivo casuale, la cellula fagocintante avrebbe degradato il batterio, già compromesso per un errore digestivo, con i sistemi di pulizia della cellula. L’osservazione è pertinente, ed è difficile rispondere dicendo che gli enzimi digestivi erano in quel momento sospesi, perché ciò avrebbe segnato una morte antecedente all’azione dell’ipotetico pre-eucariote fagogitante. Dunque, un caso fortunato. L’uguale sarebbe dovuto avvenire per il
Cianobatterio inghiottito da un batterio ameboide, e non digerito.
Le cellule fagocitanti prime (ancestrali), con solo il cloroplasto o solo il mitocondrio, dovrebbero, poi, potersi trovare da qualche parte, ma non risultano all’appello.
Tutte le alghe eucariote hanno organuli (cloroplasti - propri solo delle alghe, cioè dei vegetali -, mitocondri, nucleo provvisto di membrana, ecc.)
Si pone il problema se sia venuto prima il cloroblasto o il mitocondrio. Si è pensato di avere ritrovato degli eucarioti senza mitocondri, ma poi si è visto che avevano dei derivati dei mitocondri (mitosomi e idrognosomi).
Riguardo ai mitocondri alcuni pensano (Yacov Davidov e Eduard Jurkevitch dell’Università ebraica di Gerusalemme; James Lake, professore all’University della California a Los Angeles) a relazioni parassitarie tra procarioti - in natura questo può esistere -, considerando che i procarioti non sono capaci di fagocitosi, ma di azioni parassitarie. Le relazioni parassitarie si sarebbero sviluppate in endosimbiosi stabili, poi evolutisi in mitocondri. Ma l’evoluzione si scontra con il fatto che il parassita endosimbiotico è entrato in equilibrio con il procariota parassitato, e questo non gli può concedere una libertà di sviluppo oltre il limite endosimbiotico raggiunto.
Per le cellule vegetali e animali eucariote il nucleo si sarebbe formato da un ripiegamento interno della membrana cellulare, fino a delimitare il nucleo, non delimitato nei procarioti. In qualche modo si sarebbero formati anche gli organelli: apparato del Golgi, ribosomi, i lisosomi, il reticolo endoplasmatico, il citoscheletro. Qui, presso diversi biologi, la simbiogenesi cederebbe il posto all’evoluzione all’interno delle cellule eucariote.
Ma circa il citoscheletro c’è chi guarda al superphilum Asgard, dove sono presenti geni per proteine idonee ai citoscheletri. Anche qui con una fagocitosi fallita nella sua digestio, e tutto il resto affidato
all'immaginazione.
Il sequenziamento dei genomi
Al microscopio elettronico si aggiunse il metodo chimico di esame degli amminoacidi delle proteine e anche del DNA (1977) da parte di Walter Gilbert (1932…) e Allan Maxam (1942…). Il sequenziamento divenne più preciso con il metodo introdotto da Frederik Sanger (1918 - 2013) nel medesimo tempo (1977). Poi tutto fu ulteriormente sviluppato a partire dal 2000, con la facilitazione dell’uso dei computer. Con ciò si potevano cercare delle affinità genetiche tra batteri diversi. Esaminare il DNA circolare dei
cloroplasti e dei mitocondri e metterli in confronto con quelli dei batteri.
La composizione genomica dei cloroplasti è simile a quella dei Cianobatteri, assunti come riferimento. Diversa da quella dei nuclei.
I ribosomi dei cloroplasti sono poi simili a quelli presenti nei batteri.
I mitocontri hanno una sequenza DNA che li avvicina ai batteri, diversa da quella del nuclei delle loro rispettive cellule eucariote. Hanno ribosomi più piccoli di quelli del resto della cellula. Sono rivestiti da due membrane. La più interna ha una composizione che si avvicina a una membrana di un procariota.
Valore dei risultati
Le analisi, con tale potenza di strumenti, sono state eseguite seguendo “logiche di sperimentazione”, e hanno un valore nella ricerca delle affinità genetiche in tassonomia, tuttavia lo studio dell’informazione genetica, non chiude il problema di
come questa si traduca in un essere vivente. C’è un’indubbia relazione tra
fenotipo e genotipo, ma tale relazione, sulla quale ci si interroga, ha in sé un
determinismo che attinge alla complessità dell’unità sostanziale, vitalista, non meccanicista, di un ente biologico.
Il determinismo interno lo si può vedere dal fatto che il DNA di cellule di tessuti diversi di un organismo complesso è uguale, ma le cellule sono diversificate, obbedendo, con un processo sofisticato di attivazione e disattivazione dei geni, a un progetto di unità superiore, ben distante da una visione meccanicista.
Così, parimenti, la cellula, all’interno dell’organismo, non ha più quell’identità che avrebbe, se separata.
Neppure un organismo autonomo unicellulare ha parti (organelli) che si possano sottrarre a una unità superiore, sostanziale.
Il DNA o l’RNB sono codici che sono parte della cellula e regolano le operazioni della cellula, ma non fanno la cellula.
Un DNA isolato e messo in una coltura ricca di sostanze organiche, in parallelo all’ipotetico “brodo primordiale”, non darà mai vita a una cellula, come l’inconcludenza dell’abiogenesi dimostra.
Il percorrere la via del gene-centrismo, cioè il muoversi tra affinità, similitudini geniche, dimenticando il rapporto tra
genotipo e fenotipo, non è precisamente corretto. Le similitudini genetiche (geni ortologhi) non sono affatto uguaglianze. Inoltre un gene (geni parologhi) può codificare altra proteina. I geni ortologhi non sono delle indicazioni assolute, poiché frutto di interpretazioni. Anche una stessa proteina classificata come
omologa in due organismi non ha la stessa configurazione in percentuali di amminoacidi.
Così (Lynn Margulis), per via di tratti genetici similari, coni e bastoncelli dell’occhio, ciglia auditive delle cellule dell’orecchio interno, flagello dello spermatozoo, finiscono per avere come simbionte antenato uno
Spirochete, quanto al movimento. Spirochete inghiottito da un ente x, e poi ritenuto come endosimbionte e infine, nei millenni dei millenni, per via di simbiogenesi, giuno in qualche
trattino similare del DNA delle cellule dei vertebrati. La simbiogenesi ha punte eccessive di fantasia.
La moltiplicazione dei cloroplasti e dei mitocondri
Nella mitosi cellulare cloroplasti e mitocondri si duplicano in sincronia con la cellula, ma poi possono ancora moltiplicarsi nelle cellule.
La moltiplicazione “autonoma” dei cloroplasti nella cellula, non è affatto sgangiata dal nucleo, ma avviene, per scissione binaria, in base alla luminosità più o meno intensa e allo stato nutrizionale della pianta, Tutti sanno che una pianta posta all’ombra diminuisce la sua colorazione verde dovuta alla clorofilla dei cloroplasti.
La grande maggioranza delle proteine del cloroplasto, come per il mitocondrio, non è codificata dal suo DNA, ma da quello del nucleo della cellula, prodotte nel citosol (citoplasma), e poi immesse nel cloroplasto.
La duplicazione dei mitocondri, quella che non si attua sincronicamente con la duplicazione della cellula, avviene ugualmente per scissione binaria. Ciò risponde alle precise necessità energetiche della cellula.
Ad esempio, i mitocondri delle cellule di un atleta sono in numero maggiore, potendo aumentare con l’allenamento, di quelli di un sedentario o di un malato.
L’autonomia dei genomi circolari del mitocondrio non è però totale, perché dipende molto dal genoma del
nucleo.
La simbiogenesi seriale di Lynn Margulis
L’erudita e inarrestabile biologa Lynn Margulis, in una sua visita a Madrid nel 2006, disse all’intervistatore spagnolo Josè Manuel Nivies (ABC ciencia: 20/03/2006), circa la cellula eucariota: “Il movimento è venuto prima (spirochete). La capacità di metabolizzare l’ossigeno era seconda (mitocondri). La capacità di fotosintesi (alghe) era la terza e la quarta la produzione di proteine (archeobatteri). Ciascuna di queste capacità corrisponde, in origine, a un organismo diverso. Insieme formano le cellule eucariote di oggi”. A una domanda successiva dell’intervistatore aggiunse di intravvedere altre componenti: “Quattro sono le componenti batteriche in ogni cellula vegetale [che si alimenta con la fotosintesi]
e almeno sei in ogni cellula eterotrofica [che deve nutrirsi di composti organici]”.
Il processo presentato è una fagocitazione seriale di cellule eucariote,
necessariamente fagocitanti, dove la più grande inghiottisce, senza digestione, per un errore - per un fallimento degli enzimi -, la meno grande, per arrivare con il fissaggio, dato nei lunghi tempi, alla cellula eucariota attuale. Il caso dell’assenza di enzimi digestivi, non è
una moltitudine tale che dilaghi, da punti casuali, su tutta la terra in brevissimo tempo.
I tocchi dell’immaginazione autoevoluzionista hanno il sopravvento.
Quello che si sa, è che i reperti fossili dicono che le cellule eucariote sono apparse in tutto il mondo in brevissimo tempo (circa 1,2 miliardi di anni fa).
In un’altra intervista rilasciata il 4 marzo 2009 a Paolo Musso, professore di
Filosofia della scienza presso l’Università dell’Insubria (Il Sussidiario.net;
La Scienza in atto: 13/08/2009), a margine del Congresso internazionale a Roma organizzato dalla Pontificia Università Gregoriana, su “Biological Evolution”,
alla Margulis venne posta questa domanda: “E nel caso degli organismi superiori? Lei pensa che anche qui l’evoluzione possa spiegarsi con fenomeni di simbiosi?”. La risposa fu molto deludente riferendosi al microbiota intestinale: “Per esempio una mucca non potrebbe nemmeno digerire quello che mangia senza la collaborazione di batteri che vivono nel suo stomaco”. L’intervistatore precisò la domanda: “Ma per quanto riguarda la mutazione degli organismi in se stessi (per esempio la crescita delle dimensioni del cervello). Pensa che anche in questi casi possano essere spiegati attraverso la simbiosi?”. La risposta rivela che la sua ipotesi/teoria non reggeva più. “No, in questi casi penso che ci possano esserci altre spiegazioni (...). In ogni caso però la spiegazione non sta nelle mutazioni causali
[neodarwinismo]”. L’intervistatore pose a questo punto una domanda su come pensava il rapporto tra scienza e fede, per sondare se, in qualche modo, pensasse al creazionismo. Questa la risposta del tutto agnostica: “Ah, penso che ciascuno possa credere quello che vuole”.
Il caso di
Hatena arenicola
Pagina web consultata sull’argomento
https://schaechter.asmblog.org/schaechter/2007/05/caught_in_the_a.html
Hatena arenicola è stata scoperta nel 2000 sulla spiaggia di Isonoura in
Giappone, dopo uno tsunami, da due biologi giapponesi Noriko Okamoto e Isao
Inouve dell’Università di Tsukuba, che hanno comunicato la scoperta nel
2005. Hatena arenicola non è ancora possibile coltivarla il laboratorio, per
cui i due biologi ne hanno esaminato molti esemplari.
I due biologi hanno
segnalato il rinvenimento nel 2005, dandone descrizione scientifica nel
2006. Il nome Hatena significa “Insolito” per le sue due condizioni di vita.
Quando Hatena arenicola conduce una vita indipendente ha un’attività
predatoria di alghe. Entrando in contatto con un ceppo di alghe,
specificamente, Nephroselmis, molto abbondanti nell’habitat di Hatena
arenicola, forma con la preda una simbiosi che conduce Hatena arenicola ad
alimentarsi dei prodotti della fotosintesi dell’alga, passando così da
eterotrofo a autotrofo. La simbiosi produce modificazione nell’alga e
nell’ospite. L’alga conserva il citoplasma e il cloroplasto, che viene
ingigantito di 10 volte; i flagelli, il citoscheletro, e le membrane del
citotplasma, scompaiono; i mitocondri, l’apparato del Golgi sono degradati.
L’ospite, Hatena arenicola, acquisisce la capacità di muoversi verso la luce
(fototassi) mediante una macchia oculare algale rilevatore della luce.
Hatena arenicola perde il suo apparato nutritivo, che viene occupato dalla
macchia oculare connessa al cloroplasto.
Nella scissione per mitosi di
Hatena arenicola, ciò che resta dell’alga Nephroselmis non si divide, ma
rimane in una delle due cellule figlie, che resta di colore verde. L’altra
cellula figlia è incolore e riprende in tutto il suo carattere predatorio, e
quindi digestivo, fino a incontrare un’altra alga Nephroselmis.
Particolare
fantastico: Hatena arenicola non entra in mitosi se non dopo avere acquisito
il simbionte. Priva del simbionte non si sono mai viste divisioni.
Cosa
dire? Innanzitutto nessun input alla simbiogenesi, ma un prodigio di
endosimbiosi fatto per favorire la vita in determinate condizioni ambientali
e biologiche, con sensibilità alla luce e alla temperatura; ne è indizio che
in inverno Hatena arenicola non è ritrovabile.
Le amebe Proteus infette
Nel
1966 il microbiologo coreano Kwang W. Jeon, dell’University del Tennessee,
aveva esaminato una comunità di amebe Proteus infettate da migliaia di
batteri: 400.000 circa per ogni ameba. Quasi tutte erano morte, ma il
microbiologo selezionò quelle vive ponendole in un terreno di cultura
idoneo, il quale va visto negli esperimenti come protagonista. Dopo cinque
anni il numero dei batteri per ogni ameba era sceso a 40/42.000, essendosi
prodotta una situazione di endosimbiosi tra ameba e batteri. La notizia si
trova per la prima volta nell’ottobre del 1967 su “Journal Esperimental Cell
Research/Science Direct”. In seguito, dal 1994, Kwang W. Jeon produsse altri
articoli di ricerca sul fenomeno, su “Journal of Eukaryotic Microbiology”.
Gli esami vennero condotti usando la micrurgia e la microscopia elettronica.
Il microbiologo sottopose i batteri parassiti, poi identificati come una
varietà di Legionella, che venne chiamata Legionella Jeonii, all’azione di
antibiotici. I microbi morirono, ma, in conseguenza, anche le amebe,
lasciando intendere l’esistenza di un’endosimbiosi.
Un gruppo di Proteus
venne infettato con gli endosimbionti batterici, isolati dalle amebe
infette. I batteri evitarono l’azione enzimatica dei fagolisosomi e si
moltiplicarono più velocemente delle amebe. Nel giro di dieci generazioni
erano arrivati al numero limite di circa 42.000 per ameba, segno che le
amebe avevano attuato un sistema difensivo. I batteri erano diventati
innocui, e avevano mutato il loro moltiplicarsi. Il microscopio elettronico
rilevò la presenza di vescicole (simbiosomi), delimitate da una membrana,
che racchiudevano i batteri simbionti. Se un’ameba infetta veniva privata
dei simbionti il suo nucleo aveva anomalie irreversibili, il che voleva dire
che i batteri fornivano un’azione enzimatica ormai necessaria al nucleo.
I
simbionti, da parte loro, sopprimevano l’azione di un gene (Sams 1), che non
veniva distrutto, ma la cui espressione veniva assunta dal gene (Sams 2) con
un’azione dimezzata.
Tale azione portava al risultato che, oltre i circa
42.000 simbionti batterici, si aveva una loro moltiplicazione sotto
controllo. Tuttavia le amebe non ne traevano nessun vantaggio alimentare,
rimanendo i batteri degli endoparassiti.
Il risultato dell’esperimento di
Kwang W. Jeon è un equilibrio tra le forze dell’ameba e quelle dei batteri.
Alcune amebe, contenenti batteri endosimbionti parassiti si sono trovate in
impianti di riscaldamento centralizzati, nonché nelle acque di ambienti
ospedalieri: (Rossella Briancesco, Lucia Bonadonna: “Amebe a vita libera
nell’ambiente idrico”. Micrologia medica. Vol. 28 (3), 2013, pag 144).
[Interessano quali agenti infettivi (ma molto raramente): Acanthamoeba,
Naegleria Fowleri, Balamuthia mandrillaris, Sappina diploidea].
Le
differenziazioni delle amebe in Proteus, in Discoideis, in Echis, non sono
avvenute sotto gli occhi di Kwang W. Jeon, questo per correggere un errore
giornalistico presente nel web. Di specie di ameba ne esistono centinaia e
non derivano da un unico ceppo, ma da molti ceppi.
I nucleomorfi
Pagine web consultate sull’argomento
https://wikiita.com/nucleomorph#:~:text=I%20nucleomorfi%20sono%20piccoli%20nuclei,da%20un%20eucariota%20pi%C3%B9%20grande
https://it.m.wikipedia.org/wiki/Alga
https://en.wikipedia.org/wiki/Cryptomonad
https://www.nature.com/articles/35074092#citeas
https://www.nature.com/articles/nature11681
https://academic.oup.com/mbe/article/23/5/856/1058344
http://tolweb.org/cryptomonads
https://schaechter.asmblog.org/schaechter/2007/01/nuclear_option_1.html
Tra
tutti i gruppi di alghe, dette - accettando per comodità il termine - di
endosimbiosi secondaria, deducendo questo da tre membrane del cloroplasto, e
non da due, come nell’endosimbiosi primaria, si hanno due gruppi
(Criptomonadi e Clorarocniofite) che sono di interesse speciale per un
organulo: il nucleomorfo. Il nucleomorfo è stato intrerpretato come una
vestigia miniaturizzata del nucleo di un’alga, fagocitata da un’ameba
ancestrale, da identificare.
Tra le Criptomonadi viene segnalata l’alga
Guillarda theta, la Rhodomonas salina, la Hanusia phi, la Proteomonas
sulcata, la Cryptomonas paramecium, la Chroomonas: che è però lontanamente
imparentata.
Tra le Clorarocniofite si hanno la Bigelowiella natans, la
Clorarocniofite reptans, la Clorarocniofite Loharella.
Il nucleomorfo sarebbe in lenta fase di estinzione, fino a scomparire, poiché non compare
nei numerosi altri gruppi algali di endosimbiosi secondaria. Lo studio
genetico dei nucleomorfi ha portato all’interpretazione che quello delle
Criptomonadi procede da un’alga rossa inghiottita e non digerita per la
sospensione degli enzimi, mentre quello delle Clorarocniofite da un’alga
verde, altrettanto non digerita, secondo lo schema della simbiogenesi.
Il
nucleoformo ha tre cromosomi (sia nelle Criptomonadi, sia nelle
Clorarocniofite) avendo una sua attività residua. Il tutto presenta due
genomi minori, di tipologia procariota (mitocondri e cloroplasti); il genoma
del nucleo della cellula ospite, e il genoma accidentale del nucleomorfo. I
nucleomorfi hanno in sé un apparato genetico residuo, molto compatto, che
permette ancora la suddivisione, ma sottoposta al nucleo ospite, e rimangono
nelle cellule figlie.
Il genoma dei nucleomorfi possiede dei geni con
funzione di mantenimento: ripiegamento e degradazione delle proteine,
trascrizione e traduzione. Contiene anche geni per proteine mirate al
sistema plastidiale (cloroplasti). Esistono anche geni per l’RNA ribosomale.
Una ricerca sul trasferimento di geni dal nucleomorfo al nucleo ospite si
ritiene che richiederebbe una laboriosa ed estesa ricerca, comprendente le
alghe Criptomonadi e Clorarocniofite. Resta da sapere perché il nucleomorfo
non esista nelle altre alghe. Cosa difficile perché, oltre tutto,
bisognerebbe conoscere - al netto di ipotesi - il genoma originale della
cellula ospite (postulata dalla simbiogenesi), per valutare gli apporti.
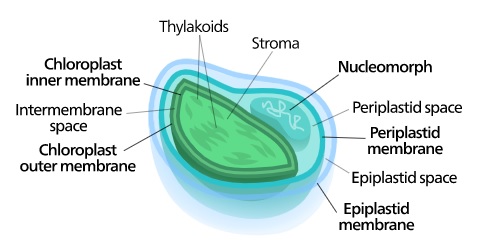
Il nucleomorfo si trova adiacente al cloroplasto che risulta avere quattro
membrane, la cui più esterna è confluente con il reticolo endoplasmatico. Ne
restano tre, e il nucleomorfo si trova tra la seconda e la terza membrana;
la prima avvolge direttamente il coloroplasto. Resta anche una minima
vestigia di citoplasma attorno al nucleomorfo. Con questo il nucleomorfo
dovrebbe essere tra la terza e la quarta menbrana e non tra la seconda e la
terza, poiché l’endosimbiosi secondaria nel suo evolversi, pur con
l’incidente del nucleomorfo, avrebbe regalato una membrana in più al
cloroplasto portandole da due a tre.
Alla fine regge il pensiero che il
nucleomorfo non sia il residuo di un’endosimbiosi secondaria, ma della
fagogitazione di un’alga non pienamente digerita, e ciò a vari gradi, come
dimostra la differenza genomica dei nucleoformi.
Le sensibili differenze dei
genomi dei nucleomorfi nei vari soggetti algali, rivela una instabilità di
fondo, che rimanda a un trauma.
Guillarda tetha, Hanusia phi, Proteomonas
sulcata, presentano una notevole variazione intracromosomica. Il genere
Chroomonas presenta nel nucleomorfo grandi genomi.
I genomi di
Hanusia phi e
di Guillarda theta, sono indistinguibili.
Anche le Clorarocniofite
presentano differenze nel genoma.
Quello che si può avanzare di fronte a
tanta complessità è che la fagocitosi di un’alga (forse non compatibile con
l’alimentazione specifica, per un’alterazione dell’habitat) da parte di una
cellula ameboide, non abbia avuto pieno successo. L’alga nel processo
digestivo dovette subire un attacco enzimatico non completo, con un trauma
delle cellule ospiti, che approdarono a una miniaturizzazione dei nuclei
algali, e a una loro variabile integrazione.
Per quel che interessa questa
indagine, invocare il nucleomorfo come segno certo dell’esistenza
dell’endosimbiosi secondaria, non ha fondamento.
Il cloroplasto con
adiacente il nucleomorfo: da Wikipedia
https://en.wikipedia.org/wiki/Nucleomorph
Un luogo comune da rigettare
Circa il luogo comune che accostarsi al mondo dell’evoluzione dei viventi porti all’agnosticismo, allo
scetticismo, al deismo, nelle loro declinazioni, va detto che agnosticismo, scetticismo, deismo, erano presenti molto prima dell’agnostico Charles Darwin (1809 - 1882) o del deista Jean Baptiste de Lamarck (1744 - 1829). Tali posizioni non emergono da chi esamina bene il vivente, se non come negazione del vero. Sono, infatti, sempre valide le parole del libro della Sapienza (13,1): “Dai beni visibili non furono capaci di riconoscere colui che è, né, esaminandone le opere, riconobbero l'artefice”. Tali parole sono ripresese nella lettera ai Romani (1, 18-21): “Infatti, dalla creazione del mondo in poi, le sue perfezioni invisibili possono essere contemplate con l’intelletto nelle opere da lui compiute, come la sua eterna potenza e divinità; essi sono dunque inescusabili”.
La scienza ha il suo oggetto specifico nel come avvengono i fenomeni, le forze in campo, le leggi che li governano, ma la scienza è fatta dall’uomo al quale è inevitabile constatare l’ordine delle cose, la loro complessità, la loro bellezzia, le leggi. Mentre lo scienziato, nel suo specifico operare, non parla di Dio, non può, nel contempo, escludere il pensiero dell’esistenza di Dio, poiché le cose sempre glielo suscitano. Non esiste un uomo, poiché uomo, che non abbia un pensiero metafisico anche embrionale, e che possa negare di averlo.
La simbiogenesi è un’ipotesi, elevata da molti a teoria per i numerosi test genetici, ma che, per i gravi vuoti, non presenta consistenza scientifica. Essa non intacca l’assunto di questo lavoro:
Evoluzione sì, ma non autoevoluzione.
E qui bisogna fermarsi e dire: Dio.
Il punto sull'evoluzione
L'evoluzionismo spiega tutto l'imponente fenomeno dell'evoluzione con quattro elementi: mutazioni casuali, nicchie ecologiche, deriva genica, simbiogenesi. Quattro elementi che non rendono per nulla ragione del grande fenomeno dell'evoluzione.
Non si ha affatto il passaggio da una specie all'altra per autoevoluzione, ma passaggi che richiedono input esterni provenienti da una causa adeguata, cioè creante: Dio.
E' giusto pensare che Dio abbia prodotto le varie specie agendo su quelle precedenti e non partendo tutte le volte dal regno minerale. Ci sono passaggi con salti
non completi (vedi Balene), ma sempre rilevanti un atto creatore su di una realtà antecedente.
Esistono gli adattamenti all'ambiente, non provenienti da errori genetici, ma da meccanismi propri della vita; vedi l'esempio dei Batteri di fronte agli antibiotici.
Esistono mutazioni che sono casuali, e questo è il solo ambito dove funziona la “teoria sintetica”.
L'evoluzione è, in conclusione, una magnifica scala offerta alla ragione per giungere a riconoscere l'esistenza di Dio trascendente e creatore.
E' bene far subentrare alla terminologia di dibattito - evoluzionismo-creazionismo - l’affermazione: “evoluzione sì, ma non autoevoluzione”.
Inserito il 7/3/2021 


|